FOURTH ADDENDUM TO THE RESEARCH AND LICENSE AGREEMENT
Exhibit 10.8
Execution Copy
FOURTH ADDENDUM
TO THE RESEARCH AND LICENSE AGREEMENT
This Fourth Addendum to Research and license Agreement (the “Fourth Addendum”) is made by and between the Technion Research and Development Foundation Ltd. (“TRDF”) and Eloxx Pharmaceuticals Ltd. (“Licensee” or “Eloxx”).
Whereas, TRDF and Eloxx are parties to a Research and License Agreement with an effective date of August 29th 2013 (the “License Agreement”), as amended on November 26th, 2013 (“First Amendment”), January 14th, 2014 (“Second Amendment”), June 9th, 2014 (“Third Amendment”), August 3rd 2014 (“First Addendum”), January 21st, 2015 (“Second Addendum”) and February 9th 2015 (“Third Addendum”) (collectively, the “Agreement”); and
Whereas, according to Section 1.26 to the License Agreement, the first Research Period has ended on September 30th, 2014 (“First Research Period”) and the First Research Plan was completed respectively (“First Research Plan”);and
Whereas, the parties desire to extend and continue the Research Period and the Research for a second year; and
Whereas, the parties desire to continue the relationship contemplated by the Agreement and therefore to further amend the Agreement as set forth herein;
NOW, THEREFORE, the parties hereby agree as follows:
| 1. | Unless otherwise defined herein, capitalized terms used in this Fourth Addendum shall have the meaning assigned thereto in the Agreement. |
| 2. | The parties wish to extend the Research Period for a second year, commencing upon the expiration of the First Research Period, on October 1st, 2014 for twelve (12) months until September 30th, 2015 (“Second Research Period”). The extension shall be on the same terms and conditions as contained in the Agreement unless otherwise agreed in this Fourth Addendum. |
| 3. | Exhibit D to the Agreement is hereby replaced with a new Exhibit D attached hereto (“Second Research Plan”). |
| 4. | The parties wish to set the terms tor the funding for the performance of the Second Research Plan during the Second Research Period, and replace section 2.2.1 of the License Agreement, as follows: |
| a) | Licensee shall fund the Research to be performed during the Second Research Period under the Second Research Plan in the total amount of fifty thousand US Dollars ($50,000) in accordance with the following schedule: |
| 1) | First installment of twenty five thousand US Dollars ($25,000) shall be paid upon signing this Fourth Addendum. |
| 2) | Second installment of twenty five thousand US Dollars ($25,000) shall be paid upon completion of the Second Research Plan and no later than October 1st 2015. |
Execution Copy
| b) | V.A.T as applicable on time of payment shall be added to each installment. |
| c) | TRDF shall issue a proper invoice for each installment. |
| 5. | Except as amended herein, all other terms and conditions of the Agreement shall remain in full force and effect. |
| ELOXX PHARMACEUTICALS LTD. DEVELOPMENT | THE TECHNION RESEARCH & FOUNDATION LTD. | |||
| By: | /s/ Xxxxxx Xxxxxx |
By: | /s/ Mano Medalsi |
|||||||
| Name: Xxxxxx Xxxxxx | Name: Mano Medalsi, CPA | |||||||||
| Title: CEO | Title: | |||||||||
| Date: 24/4/15 | Date: 29/4/15 | |||||||||
Exhibit D
Submitted to Eloxx Pharmaceuticals LTD
Research Plan
(Continuation for the second year 1.10.2014-30.092015)
Title
Development of Aminoglycose-Based Drug for Treatment of Human Genetic Diseases and Many Forms of Cancer Caused by Nonsense Mutations
I. Brief Summary of the First Year’s Research
The two specific aims were: To provide Eloxx with all the important lead compounds developed by us in appropriate quantities (as agreed and signed) for the validation of our previously published data, along with the detailed protocols of the synthesis and biological evaluations performed in our labs (Aim #1); Further modifications of the developed leads and/or development of new lead structures for improved efficacy as potential drugs (Aim #2). The main achievements include:
(1) We successfully synthesized all the lead compounds, provided them to Eloxx, and the company has successfully validated with these compounds the entire biological and toxicity data previously reported by us.
(2) We have generated a new series of N1-alkyl derivatives on the disaccharide scaffold and demonstrated their improved activity in comparison to parent compounds. Encouraged, the best disaccharide derivative NB146 was further converted to the corresponding trisaccharide derivative, NB147. Both these new leads (NB146 and NB147) were provided to Eloxx for further assessment their biological tests.
(3) By developing a simple and robust synthetic scheme for the modification of the developed leads as poly-ester derivatives, we have produced two such new derivatives Bz-G418 (a prototype structure for the proof-of-concept) and Bz-NB124 (along with their parent structures) and provided them to Eloxx for further assessment their biological tests.
In addition, two workshops have organized by the PI during this grant period with Eloxx together with a professional in CF, Xxxx X. Xxxx (UAB, USA) and the professional in Hurler syndrome, Xxxx. X. Xxxxxxx (UAB, USA) to critically discuss our recent developments and future progresses.
II. Proposed Working Plan for the Second Year Research
Specific Aims
Aim # 1: Rational Design and Synthesis of New Readthrough Drugs. Our design principles integrate the insights from the first year’s achievements in our lab along with the insights in other labs in order to construct new classes of compounds by a structure-based approach. Several sets of structurally distinct chemical compounds will be included in the initial library and promising lead compounds will be further refined for better cell permeability and prolonged in-vivo action. The synthesis will use state-of-the-art strategies for the assembly of complex carbohydrate, along with the convenient up-to-date analytical techniques.
Aim #2: Chemical Modification of NBs for Improved Cell Permeability and Oral Bioavailability. We will continue the project from the first year on the polyester derivatives of NBs and this strategy will be further expanded with polyamide derivatives and with the special modifications of NBs as pro-drugs for better blood-brain-barrier (BBB) permeability.
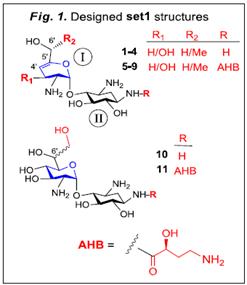
Aim #1- (a) rational design of pseudo-disaccharide scaffolds: Based on our previous results with development of lead structures of NB-series, we propose to modify these compounds as well as to extend our study to more diverse structures with potentially improved suppression activity and lower toxicity. We will probe the pseudo-disaccharides of set1 (Fig. 1) as the minimum basic structures. The rationale in selecting them is based on our observations with previous leads exhibiting C6’-OH, and that further modifications by inserting either unsaturation at ring I (glucosamine ring, compounds 1-9) or the 6’,7’-diol (compounds 10-11) are expected to improve their specificity
Timor Baasov

towards the cytoplasmic A-site, leading to enhanced readthrough and reduced
toxicity. This expectation is supported by: (i) the aminoglycoside (AG) sisomicin that exhibits a similar unsaturated ring between C4’- C5’ at ring I displays better antibacterial activity than similar
4,6-disubstituted AGs with a saturated ring I, like gentamicin, tobramycin and amikacin. The superior antibacterial activity of sisomicin has been attributed to its specific binding mode1: while a typical saturated ring I with a chair conformation stacks on G1491 through CH-p interactions, the unsaturated ring I of sisomicin stacks with a
partially planar conformation through p -p interactions with G1491 and achieves a closer fit within the binding pocket; (ii) while sisomicin is active
only against bacteria, 6’-hydroxysisomicin possesses promising antiprotozoal activity2: a comparison of stacking interactions between G1491 and ring I of 6’-hydroxysisomicin and
geneticin in the protozoal A-site clearly favors the former (Fig. 2A). 6’-hydroxysisomicin can adopt a lower energy conformation within the protozoal A-site,
leading to the entropically favorable burying of the hydrophobic region of ring I with optimized p-p stacking on G1491. (iii) Our recently solved crystal
structure of G418 bound to a model of the Leishmania ribosomal A-site supported the above notions3: the ring I of G418 is fixed into the Leishmania A-site due to a stable pseudo pair with G1408 (Fig. 2B) and the additional H-bonds O3’
 and
and
 on the opposite site of the pseudo pair (Fig. 2C), leading to an unfavorable stacking of ring I with the A1491. [Note that one of the major differences between the protozoal and Leishmanial A-sites is the nucleotide at position 1491 (E. coli numbering): G1491 in most protozoa and A1491 in Leishmania; therefore the Leishmanial ribosome shares greatest similarity with human ribosomes, with only a single
nucleotide differentiating the AGs putative binding sites (U1409 in Leishmania vs. C1409 in human]. Therefore, the deletion of only C4’-XX (0-0, X0xXX, Fig 1) with simultaneous introduction of C4’-C5’ double bond, can make ring I relatively free to move within the binding pocket and form
stronger n-n stacking with A1491.
on the opposite site of the pseudo pair (Fig. 2C), leading to an unfavorable stacking of ring I with the A1491. [Note that one of the major differences between the protozoal and Leishmanial A-sites is the nucleotide at position 1491 (E. coli numbering): G1491 in most protozoa and A1491 in Leishmania; therefore the Leishmanial ribosome shares greatest similarity with human ribosomes, with only a single
nucleotide differentiating the AGs putative binding sites (U1409 in Leishmania vs. C1409 in human]. Therefore, the deletion of only C4’-XX (0-0, X0xXX, Fig 1) with simultaneous introduction of C4’-C5’ double bond, can make ring I relatively free to move within the binding pocket and form
stronger n-n stacking with A1491.
The rationale for the selective attachment of (S)-4-amino-2-hydroxybutanoyl (AHB) at the N1 position of set1 (R=AHB) is based on the highly positive impact of this group (for example in NB84) compared to their parent structures (for example NB74), respectively4-8. Therefore, we will continue this approach in new designs. Finally, in all set1 structures we preserved the C6’-OH function as a crucial motif for favorable H-bond interactions between ring I of AG and G1408 (Fig. 2B). We anticipate that the deletion of C4’-OH or C3’,C4’-hydroxyls with a simultaneous introduction of unsaturation on ring I will make the ring relatively “free” to move within the binding pocket for better pseudo-pair interaction with G1408 and improved p-p stacking with A1491 (Fig. 2C). The recent comparative x-ray data of G418 and 6’-hydroxysisomicin in complex with the bacterial and protozoal A-sites support this expectation1-2 (Fig. 2A). In order to form a pseudo pair with 6’- hydroxysisomicin, the G1408 residue in the protozoal A-site shifts toward the deep/major groove compared with the position of A1408 in bacterial A-site, while ring I rotates approximately 13º around O4- C1’ glycosidic bond between rings I and II; allowing the formation of a stable pseudo-pair between the ring I of 6’- hydroxysisomicin and G1408, along with the simultaneous strong p-p stacking of ring I with G1491.
Timor Baasov
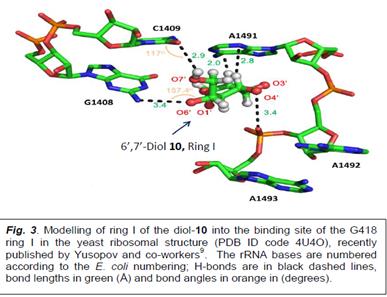
While these data support the likelihood that the compounds 1-9 of set1 structures (Fig. 1) with 6’-OH and unsaturation at ring I may perform better than previous leads with the saturated ring I, we want to introduce additional structural motifs at the C6’- region that would further increase this expectation. We plan to replace the CH2OH (and/or CH(CH3)OH) at 6’-position with the vicinal 6’,7’-diol (compounds 10-11, Fig.1). The replacement of C6’-OH with 6’,7’-diol is expected to generate stronger H-bond interactions of ring I with A site residues G1408 and C1409 resulting in better pseudo-disaccharide scaffold of set1 (Fig. 3). Following important points in Fig. 3 are of note. First, unlike our solved structure of G418 bound to Leishmania A-site (Fig. 2B and 2C) in which ring I of G418 makes two H-bonds with G1408, the structure of G418 bound to the entire yeast ribosome recently reported by Xxxxxxx and co-workers9, shows only one H-bond between O6’ and G1408 nitrogen (Fig. 3). Modelling ring I of the 6’,7’-diol 10 onto this structure of Yusopov, clearly shows that O7’-OH can make additional H-bond with the oxygen of C1409.
To test this issue, we initially synthesized compound 10 (Fig. 1) as a single diastereomer at C6’ with unknown configuration (the complete assignment of the absolute configuration of 10 at C6’, along with the synthesis of its C6’- distereomer are underway). Preliminary in-vitro suppression tests demonstrated that 10 exhibits superior readthrough to that of 12, and paromamine (Fig. 4). Interestingly, installation of 6’,7’-diol on the paromamine scaffold (10) dramatically increases its in-vitro readthrough activ ity. It is of note that the observed activity increase in 10 is far higher than the impact of the chiral (R)-6’- methyl group in 12, which was to date our most favorable pharmacophore at C6’- regon7,8. With compound 10 we are already one step ahead! Further installation of chiral (R)-6’-methyl group and/or N1-AHB group on compound 10 will likely provide additional improvement of this performance, and these potentials will be tested in the subsequent steps.
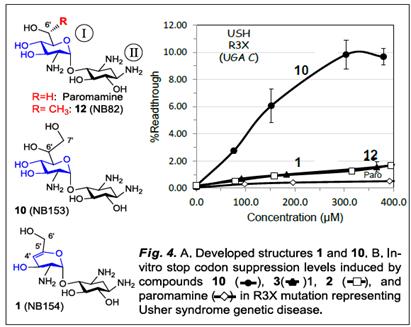
In addition to 10 we also synthesized the simplest representative of set1 structures, compound 1 (R=R1=R2=H, Figs. 1 and 2). Preliminary comparative in-vitro suppression tests demonstrated that 1 exhibits superior readthrough to that of its parent paromamine and is very similar to that of compound 12 (Fig. 4). These data suggest that the anticipated p-p stacking interaction of the unsaturated ring I of 1 with A1491 is much stronger than the CH-p interactions of the saturated ring I of paromamine with A1491. The data in Fig. 4 also suggests that the advantage of such stacking interactions in 1 (paromamineg1) is similar to that of (R)-6’-methyl group in 12 (paromamineg12), and thus identifies unsaturation at ring I as a potential new pharmacophore.
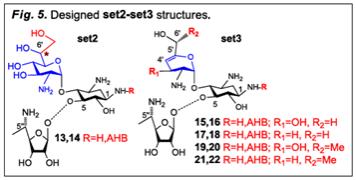
Aim #1- b) rational design of pseudo-trisaccharide scaffolds: It should be noted that the set1 structures (Fig. 1) are not expected to possess significant PTC suppression activity (Fig. 4); rather they will be used as scaffolds for the construction of new efficacious structures by attaching various sugar and/or acyclic appendages. For a systematic study, the same modifications will be performed on all pairs of set1, with the aim to reach the best lead compound. As a ring III we have developed a new structure of (S)- 5’’-amino-5’’-methyl-D-ribose, and its advantage over 5”-aminoribose was evident in our previous leads7,8. Therefore, we will use this novel ring III for the generation of new pseudo-trisaccharides (set2-set3, Fig. 5). Encouraged by the preliminary data obtained with 10 (Fig. 4), we will scrutinize the stereochemistry at C6’ of 10 and will assemble the corresponding pseudo-
6
Timor Baasov
trisaccharides 13 and 14 (set2, Fig. 5), along with all the variations of pseudo-trisaccharides with unsuterated ring I (compounds 15-22, set3, Fig. 5). In addition, based on the observed strong impact of the vicinal 6’,7’-diol in 10 (Fig. 4), this variable (6’,7’-diol) will further be added to the planned pseudo-trisaccharides of set3 structures in Fig. 5, to yield additional sets of compounds incorporating the vicinal 6’,7’-diol and unsaturation at ring I.
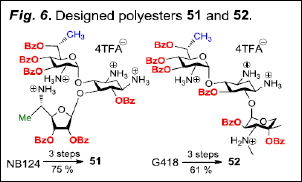
Aim #2. a) Modification of lead structures for improved cell permeability and oral bioavailability: Because AGs are highly charged, water soluble compounds, they have poor intestinal absorption and are usually injected. In addition, AGs are short-lived in the circulatory system, rapidly eliminated by glomerular filtration in the kidney. They also exhibit poor permeability into eukaryotic cells, which requires higher dosages that often cause harmful side effects that limit their use in therapy. In order to overcome these limitations we designed different sets of compounds. One of such sets (Fig. 6) considers that all hydroxyls of the AG are replaced by esters. The rationale is to increase the lipophilicity and cell-permeability of the drugs, improve in-vivo half-life and oral bioavailability. These changes should provide important therapeutic benefit over treatment with the initial lead alone.
The suggested modifications on NB124 and G418 (Fig. 6) to yield the corresponding per-benzoate esters (OBz, 51 and 52, respectively) are based on the following observations. (i) In our most recent comparative study of our lead compounds using an extended repertoire of CF cell lines, reporters, assays and animal models, comp. NB124 was able to most effectively rescue CFTR function, was superior to gentamicin, exhibited favorable pharmacokinetics, and was less cytotoxic than gentamicin in the explant model of ototoxicity10; (ii) the benzoate esters 51 and 52 are chemically more stable than the corresponding simple alkyl asters (like acetate, isobutirate, isopropionate); (iii) the poly-esters 51 and 52 can easily be prepared in good overall yields from the corresponding AGs in three steps (Boc2O, Et3N, H2O/MeOH; BzCl, Py, 4DMAP; TRA, CH2Cl2); (iv) to test our hypothesis we initially used 52 as a prototype derived from commercial G418.
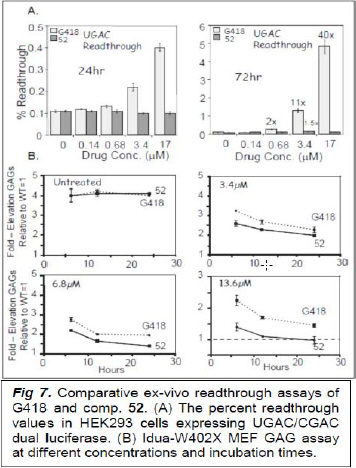
Preliminary tests of 52 against its parent G418 demonstrated that 52 lacks any readthrough activity both in cell free (see the data in the 1st year research report) assays and HEK293 cells stably expressing UGAC/CGAC dual luciferase (Fig. 7A) even when the incubation time was extended up to 72 hrs. Interestingly, when we tested the antibacterial activities of G418 and 52, we found that while the activity of 52 was similar to that of G418 against various G- and G+ WT bacteria, the MIC values of 52 were significantly lower than that of G418 in bacterial strains harboring plasmids of AG resistance determinant enzymes (see the data in the 1st year research report). Encouraged, we argued that HEK293 cells, being engineered cell lines, might not contain the required esterases to hydrolyze the benzoate esters in 52. Indeed, using MEFs derived from homozygous Idua-W402X mice11, we found that the activity of 52 significantly exceeds that of G418 at all incubation times and concentrations tested. At a dose of only 13.6 micromolar, treated cells had already reached the glycosaminoglycan (GAG) levels found in WT cells (Fig. 7B). These encouraging data with 52 prompted us to prepare compound 51 (see the data in the 1st year research report), which is currently under similar tests in Idua-W402X MEFs (by our collaborator X. Xxxxxxx at Univ. of Alabama, Birmingham) and CF cell lines (primary human bronchial epithelial, HBE cells, G542X/delF508, X. Xxxx at the UAB). In-vivo tests in the Idua-W402X mice (X. Xxxxxxx) will follow these experiments.
7
Timor Baasov

Aim #2 b) Modifications for improved capacity to pass the blood-brain barrier (BBB). While we have been successful in identifying promising nonsense suppression agents (NB84) for the Idua-W402X mutation and partially alleviating the primary GAG storage defect associated with MPS I-H within a variety of tissues (including brain), we have been unable to completely restore GAG levels in the mouse model, even in a long-term, 28-week drug-treatment12,13. Even though the newly proposed sets of compounds might prove better than the previously examined leads (especially the structures with elevated lipophilicity like 51) for treatment of MPS I-H, we propose to also test a brain-targeted chemical delivery system (CDS)14 which is expected to produce a more robust therapeutic response. The approach is based on the attachment of a dihydropyridine moiety as a carrier for delivering drugs through the BBB (Fig. 8). This site-selective delivery will reduce the exposure of the body to high drug levels and consequently increase their therapeutic index. Since NB84 has already been proven to significantly moderate disease progression in-vivo in our MPS I-H mouse xxxxx ,00 we will now synthesize the new derivative of NB84, the compound NB84-CDS (Fig. 11). We (our collaborator at UAB, X. Xxxxxxx) will then determine whether delivery of NB84-CDS through the BBB is better than NB84 in a long term study using established biochemical and immunohistochemical assays as previously described12.
REFERENCES
| (1) | Kondo, J., Koganei, M., and Kasahara, T. (2012) Crystal Structure and Specific Binding Mode of Sisomicin to the Bacterial Ribosomal Decoding Site. ACS Med. Chem. Lett. 3,9, 741-744. |
| (2) | Kondo, J., Koganei, M., Maianti, J. P., Xx, X. X., and Xxxxxxxxx, S. (2013) Crystal Structures of a Bioactive 6’-Hydroxy Variant of Sisomicin Bound to the Bacterial and Protozoal Ribosomal Decoding Sites. ChemMedChem 8, 733–739. |
| (3) | Shalev, M., Kondo, J., Kopelyanskiy, D., Xxxxx, X. X., Adir, N., and Baasov, T. (2013) Identification of the molecular attributes required for aminoglycoside activity against Leishmania. Proc. Natl. Acad. Sci. U. S. A. 110, 13333–13338. |
| (4) | Xxxxxxxx, I., Xxxxxx-Xxxxxx, A., Xxxxxxxxxxx, M., Xxxxxxxx, V., Hainrichson, M., Chen, F., Xxxxxxx, J., Xxxxx, X. X., Xxx-Xxxxx, T., and Baasov, T. (2009) Development of novel aminoglycoside (NB54) with reduced toxicity and enhanced suppression of disease-causing premature stop mutations. J. Med. Chem. 52, 2836–2845. |
8
Timor Baasov
| (5) | Xxxxxxxx, I., Xxxxxx, D., Xxxxxxx, B., Hainrichson, M., Xxxxxxxx, V., and Baasov, T. (2010) Repairing faulty genes by aminoglycosides: development of new derivatives of geneticin (G418) with enhanced suppression of diseases-causing nonsense mutations. Bioorg. Med. Chem. 18, 3735–3746. |
| (6) | Xxxxxx-Xxxxxx, A., Xxxxxxxx, I., Xxxxx, X. X., Baasov, T., and Xxx-Xxxxx, T. (2007) In vitro and ex vivo suppression by aminoglycosides of PCDH15 nonsense mutations underlying type 1 Xxxxx syndrome. Hum. Genet. 122, 373–381. |
| (7) | Xxxxxxxxx Xxxx-Xxxxxx, D., Xxxxxxxx, V. and Baasov, T., J. (2011) Repairing faulty genes by aminoglycosides: Identification of new pharmacophore with enhanced suppression of diseases-causing nonsense mutations. Med Chem Comm 2, 165–171. |
| (8) | Xxxxxxxxx, J., Xxxx-Xxxxxx, D., Xxxxxxx, E., Xxxxxxx, K., Shavit, M., Xxxxxxxx, V., and Baasov, T. (2012) Increased selectivity toward cytoplasmic versus mitochondrial ribosome confers improved efficiency of synthetic aminoglycosides in fixing damaged genes: A strategy for treatment of genetic diseases caused by nonsense mutations. J. Med. Chem. 55, 10630– 10643. |
| (9) | De Loubresse, N. G., Prokhorova, I., Xxxxxxxx, W., Rodnina, M. V., Xxxxxxxx, G., and Xxxxxxx, M. (2014) Structural basis for the inhibition of the eukaryotic ribosome. Nature 513, 517-522. |
| (10) | Xxxx, G., Dai, Y., Du, M., Xxxxxxxx, V., Xxxxxxxxx, J., Xxxxxx, X. X., Baasov, T., Xxxxxxx, X. X., and Keeling, K. M. (2014) Long-term nonsense suppression therapy moderates MPS I-H disease progression. Mol. Genet. Metab. 111, 374-381. |
| (11) | Keeling, K. M., Wang, D., Dai, Y., Xxxxxxxxx, S., Xxxxxx, B., Xxxxx, J., Xxxxxxxx, V., Xxxxxxxxx, J., Velu, S. E., Baasov, T., and Xxxxxxx, X. X. (2013) Attenuation of Nonsense-Mediated mRNA Decay Enhances In Vivo Nonsense Suppression. PLoS One 8, 4, e60478. |
| (12) | Xxxx, G., Dai, Y., Du, M., Xxxxxxxx, V., Xxxxxxxxx, J., Xxxxxx, X. X., Baasov, T., Xxxxxxx, X. X., and Keeling, K. M. (2014) Long-term nonsense suppression therapy moderates MPS I-H disease progression. Mol. Genet. Metab. 111, 374-381. |
| (13) | Keeling, K. M., Wang, D., Dai, Y., Xxxxxxxxx, S., Xxxxxx, B., Xxxxx, J., Xxxxxxxx, V., Xxxxxxxxx, J., Velu, S. E., Baasov, T., and Xxxxxxx, X. X. (2013) Attenuation of Nonsense-Mediated mRNA Decay Enhances In Vivo Nonsense Suppression. PLoS One 8, 4, e60478. |
| (14) | Xxxxx, N., and Xxxxxxxx, P. (2008) Retrometabolic drug design: Principles and recent developments. Pure Appl. Chem. 80, 1669-1682. |
III. Proposed Budget (in US Dollars) and Time Frame
| The proposed task | Time Frame | Budget Requested |
Remarks | Notes | ||||||||
| Aim # 1 |
Establishment of config. In 10 and synthesis of 13. | 6-12 months |
$ | 15,000 | One postdoc/student dedicated. | PI T. Baasov | ||||||
| Synthesis of 15 and preliminary biochem. Analysis of 10,13 and 15. | 6-12 months |
$ | 15,000 | One postdoc/student dedicated. | ||||||||
| Total Aim #1: | $ | 30,000 | ||||||||||
|
|
|
|
||||||||||
| Aim # 2a |
Synthesis of NB84-esters and NB124-esters | 8-12 months |
$ | 10,000 | One postdoc/student dedicated. | |||||||
| Aim # 2b |
Synthesis of NB84-CDS and its Preliminary biochemical analysis | 6-12 months |
$ | 10,000 | One postdoc/student dedicated. | |||||||
| Total Aim #2: | $ | 20,000 | ||||||||||
|
|
|
|
||||||||||
| Grand total for the Aims # 1&2: | $ | 50,000 | ||||||||||
|
|
|
|
||||||||||
9
Timor Baasov
NOTE the budget does not include ex vivo (MEFs) and in vivo studies required for the project completion and only can be done with additional funding of collaborators labs (Xxxxxxx and Xxxx at UAB)!
IV. Detailed Budget (in US Dollars)
Personnel:
| Role in project | % Time | Salary | ||||||||||
| 1. |
Lab. Assistant |
Lab. Assistant |
50 | 12,000 | ||||||||
| 2. |
Postdoctorant |
Researcher |
50 | 12,000 | ||||||||
|
|
|
|||||||||||
| Total: |
24,000 | |||||||||||
Supplies:
| 1. Chemicals, absolute & deuterated solvents |
18,000 | |||
| 2. Lab. equipment, glassware, plastic xxxx |
2,000 | |||
| 3. Biochemicals |
2,000 | |||
| 4. In vitro and ex vivo assay kits |
2,000 | |||
| 5. Chromatography material for various purifications |
1,000 | |||
| 6. NMR and Mass Spectrometry tests |
1,000 | |||
|
|
|
|||
| Total: |
26,000 | |||
|
|
|
|||
| Grand Total: |
$ | 50,000 |
Budget Justification:
The requests for laboratory assistant and postdoctorant for the duration of the grant period are in recognition of the amount of work required in this project. Considerable effort will be expended in the syntheses of various lead compounds discussed in the proposal, their structure determination and analysis, assays for their activity. The request for materials, supplies, and chemicals/biochemicals is an important part for a successful development and completion of the project.
V. Investigators’ Curriculum Vitae
| Surname: | Baasov |
First name: | Timor | |||||
| Birthdate: | January 3, 1954 |
(a) Education Background
| From-To |
Institution |
Area of specialization |
Degree | |||
| 1981-1986 |
Weizmann Institute of Science | Chemistry | Ph. D. | |||
| 1977-1979 |
Tel-Aviv University | Chemistry | M. Sc. | |||
| 1975-1977 |
Tel-Aviv University | Chemistry | B. Sc. |
Major research interest: Carbohydrate chemistry, Bioorganic and medicinal chemistry, Drug design and development, Rational design of substrate and inhibitors, Mechanistic enzymology.
(b) Employment
| From-To |
Institution |
Research area |
Title | |||
| 2004- |
Technion | Bioorganic Chemistry | Professor | |||
| 3/1998-8/1998 |
The Scripps Research Institute, Cal | Bioorganic Chemistry | Visiting Prof. |
10
Timor Baasov
| 1998-2003 |
Technion | Bioorganic Chemistry | Assoc. Prof. | |||
| 1990-1998 |
Technion | Bioorganic Chemistry | Senior Lecturer | |||
| 1988-1990 |
Technion | Bioorganic Chemistry | Lecturer | |||
| 1986-1988 |
Harvard University | Bioorganic Chemistry | Post-Doct. Res. |
VI. T. Baasov - List of Publications last three years (2011-2014)
| 1. | X. Xxxxxxx, X. Xxxxxxxx, X. Xxxxxx, X. Xxxxxxx, X. Xxxxxxxx, X. Xxxxxxxx, X. Xxxxxx, X. Xxxxxx. Readthrough of Nonsense Mutations in Rett Syndrome: Evaluation of novel aminoglycosides and generation of a new mouse model. Journal Molecular Medicine, 89, 389-398, (2011). |
| 2. | X. Xxxxxxxxx, X. Xxxx-Xxxxxx, X. Xxxxxxxx, X. Xxxxxx. Repairing faulty genes by aminoglycosides: Identification of new pharmacophore with enhanced suppression of diseases-causing nonsense mutations. Medicinal Chemistry Communications, 2, 165-171 (2011). |
| 3. | X.X. Xxxx, X.X. Xxxx, X. Xxxxxx, X.Xxxxxx, X. Xxxxx, X. Xxxx;xx-Xxxxxxx, X. Xxxxxxxx, X. Xxxxxxxx, X. Xxxxx, X. Xxxxxxxxxx, X. Xxxxxx, X.X. Xxxxxxx. Suppression of CFTR Premature Termination Codons and Rescue of CFTR Protein and Function by the Synthetic Xxxxxxxxxxxxxx XX00. Journal Molecular Medicine (Berl), 89, 1149-1154 (2011). |
| 4. | X. Xxxxxxx, B. Xxx Xxxx, X. Xxxxxxxx, X. Xxxxxxxx, X. X. Xxxxx, X. Xxxxxxxxx, G. Rechavi, T. Baasov, E. Gak. Ex Vivo Treatment with a Novel Synthetic Aminoglycoside NB54 in Primary Fibroblasts from Rett Syndrome Patients Suppresses MECP2 Nonsense Mutations. PLoS ONE, 6 (6), e20733 (2011). |
| 5. | H-L. X. Xxx, C-C. Xxxx, X. Xxxxxx, Y. Xxx, X. X. Xxxxxxxxx. Post-transcriptionally Regulated Expression System in Human Xenogeneic Transplantation Models. Molecular Therapy, 19(9), 1645-1655 (2011). |
| 6. | X. Xxxx , X. Xxxxxxxx, X. Xxxxxxxxx, X. Xxxxxx, S-C. Li, Y-T Li, X.X. Xxxxxxx, X.X. Xxxxxxx. The designer aminoglycoside NB84 significantly reduces glycosaminoglycan accumulation associated with MPS I-H in the Idua-W392X mouse. Molecular Genetics and Metabolism 105, 116-125 (2012). |
| 7. | X. Xxxxxxxx, X. Xxxxxxxx, X. Xxxxxx, X. Xxxxxxxx, M. van Wyk, T. Baasov, X. Xxxxxxx, and X. Xxxxx-Xxxxxxx. A comparative evaluation of XX00, XX00 and PTC124 in translational read-through efficacy for treatment of an USH1C nonsense mutation. EMBO Molecular Medicine, 4, 1-14, (2012). |
| 8. | X. Xxxxxxxxx, X. Xxxx-Xxxxxx, X. Xxxxxxx, X. Xxxxxxx, X. Xxxxxx, X. Xxxxxxxx X. Xxxxxx. Increased Selectivity toward Cytoplasmic versus Mitochondrial Ribosome Confers Improved Efficiency of Synthetic Aminoglycosides in Fixing Damaged Genes: A Strategy for Treatment of Genetic Diseases Caused by Nonsense Mutations. J. Med. Chem. 55(23), 10630-10643 (2012). |
| 9. | M. Schalev, X. Xxxxxxxxx, X. Xxxxxx, X. Xxxxxxxx, X. Xxxxx-Xxxxxxxxx, X. Xxxxxx. Development of generic immunoassay for the detection of a series of aminoglycosides with 6’-OH group for the treatment of genetic diseases in biological samples. Journal of pharmaceutical and biomedical analysis. 75, 33-40 (2013). |
| 10. | X.X. Xxxxxxx, X. Xxxx, X. Xxx, X. Xxxxxxxxx, X. Xxxxxx, X. Xxxxx; X. Xxxxxxxx, X. Xxxxxxxxx, X.X. Xxxx, X. Xxxxxx, X.X. Xxxxxxx. Attenuation of Nonsense-Mediated mRNA Decay Enhances In Vivo Nonsense Suppression. PLoS ONE 8 (4), e60478 (2013). |
| 11. | M. Schalev, X. Xxxxx, D. Kopelyanskiy, X.X. Xxxxx, X. Xxxx, X. Xxxxxx. Identification of the molecular attributes required for Aminoglycoside activity against Leishmania. PNAS 110 (33), 13333-13338 (2013). |
| 12. | X. Xxxxx, X. Xxxxxxxxx, X. Xxxxxx, X. Xxxxxxxxx-Xxxxxxxx, X. Xxxxxxxxxxx,, X. Xxxxxxxx, X. Xxxxxx, X.X. Xxxxxxx, X.X. Xxxxxx. Am inoglycoside-Induced Premature Stop Codon Read-Through of Mucopolysaccharidosis Type I Patients Q70X and W402X Mutations in Cultured Cells. Journal of Inherited Metabolic Disease Reports. 13, 139-147 (2014) |
| 13. | X. Xue, X. Xxxxxx, X.X. Xxxx, X. Xxxxxx, M. Du, L. A. Jackson, Y. Dai, X. Xxxxxxxx, X. Xxxxxx, X. Xxxx, X. Xxxxxxx, X. Xxxxxxx, X. Xxxxxx, X. Xxxx, X. X. Xxxxxxx, X.X. Xxxx. Synthetic Aminoglycosides Efficiently Suppress CFTR Nonsense Mutations and Are Enhanced by Ivacaftor. Am. J. Respir. Cell Mol. Biol. 50 (4), 805- 816 (2014). |
| 14. | X. Xxxxxxx, X. Xxxxxxxx, X. Xxx, X. Xxxxxxx, E. G. Meyron-Xxxxx, X. Xxx-Xxxxxxx, X. Xxxxxxx, X. Xxxxxx. Designer aminoglycosides that selectively inhibit cytoplasmic rather than mitochondrial ribosomes show decreased ototoxicity: a strategy for the treatment of genetic diseases. J. Biol. Chem. 289(4), 2318-2330 (2014). |
| 15. | X. Xxxx, Y. Dai, M. Du, X. Xxxxxxxx, X. Xxxxxxxxx, X.X. Xxxxxx, X. Xxxxxx, X.X. Xxxxxxx, X.X. Xxxxxxx. Long-term nonsense suppression therapy with NB84 moderates MPS IH disease progression. Molec. Genet. Metabol. 111, 374-381 (2014). |
11
Timor Baasov
| 16. | X. Xxxxxx, T. Baasov. When Proteins Start to Make Sense: Fine-tuning of Aminoglycosides for PTC Suppression Therapy. Med. Chem. Commun. 5, 1092-1105 (2014). Invited Review Perspective Article |
Chapters in Books and other Publications
| 1. | X. Xxxxxx, X. Xxx, X. Xxxxxxx, X. Xxxx, X. Xxxxxx, X. Xxxxxxx, X. Xxxxxx, X. Xxxxxxxx, X. Xxxxxxx, X. Xxxx. Use of transepithelial conductance as a screening technique for identification of drugs that promote readthrough of premature stop codons. Pediatric Pulmonology 48, page 232 (supplement 36; meeting abstract). ISSN: 8755-6863 (2013). |
| 2. | X. Xxxxx-Xxxxxxx, T. Baasov, X. Xxxxxxx. Therapy strategies for Xxxxx syndrome Type 1C in the retina. Advances in experimental medicine and biology. Vol. 801, pp. 741-747 (2014). |
| 3. | T. Baasov, X. Xxxxxxx. Foreword-The 17th European Carbohydrate Symposium-EuroCarb17. Carbohydrate Research 389, 1 (2014). (Guest Editor of the special issue.) |
| 4. | X. Xxxxxxx-Xxxxxxxxx, T. Baasov. Editorial – Carbohydrates Themed Issue. Med. Chem. Commun., 5, 1010-1013 (2014). (Guest Editor of the special issue.) |
12

